Anyone who has ever watched a close football game has seen this scenario. Time is running out, and the team that’s behind has one play to get the ball all the way down the field for a touchdown. While a long pass thrown is an option, the chances of success are slim; such a play is referred to as a “Hail Mary” for a reason. What often happens instead is the quarterback will pass the ball to a nearby player who, in turn, will pass the ball to a teammate before getting tackled. What ensues is a series of short passes from player to player. The “trick” to this play is that all the passes after the initial forward pass, by rule, must go backwards. Players try to stay close to one another to keep the passes short. Most of these players are not trained to throw the ball, and care must be taken to keep the passes going backward while the players are moving forward. Players also need to block defenders trying to knock the ball or its carrier to the ground. As exciting as these plays are, inevitably, they come up short. There are just too many moving parts, and so much can go wrong.
The Improbable Electron Transport Chain
A biochemical process that is more improbable than a game-winning trick play constantly takes place in the inner membrane of mitochondria. This process—called cellular respiration—is a set of metabolic reactions that produces the energy that the cell needs. The electron transport chain (ETC) is one portion of that process. The ETC consists of a series of proteins embedded in the inner mitochondrial membrane. The whole enterprise of the ETC would not be possible if not for a handful of metallic elements buried deep within the complex structure of the proteins of the ETC. These proteins “pass” electrons down the chain from metallic element to metallic element in an exquisitely fine-tuned process that I see as a reflection of the Creator’s handiwork.
The source of the electrons in the ETC are the molecules NADH and FADH2, which are produced from the citric acid cycle. These electrons ultimately react with H+ ions and molecular oxygen to form H2O, but the beginning and end of the process is separated not only by physical space, but also by an energy or potential difference. Electrons don’t move from metal center to metal center unless it is energetically favored. Furthermore, electrons can’t just wander through the cell over long distances without reacting with something.
A Biochemical Playbook
In biochemical systems, the behavior of these electron transfers is described by Marcus theory,1 and for decades biochemists have been investigating exactly how the mechanism works, and how such a complex system ended up in cells. While Marcus theory is a complex model describing electron transfer, it really boils down to two essential ideas: (1) electron transfer must be energetically favorable between two metal centers, and (2) the proteins that house those metallic elements need to be really close together to ensure sufficient kinetics or speed. This is not too different from our crazy football play. To increase the chances of success, the ball has to be thrown in the right direction as a series of short passes.
In protein Complex I of the ETC, NADH undergoes oxidation to NAD+ and H+. The molecule that initially accepts the hydrogen ions and electrons is an organic molecule called FMN (flavin mononucleotide). A series of “hops” by the electrons between clusters of iron and sulfur atoms follows this reaction. These iron-sulfur clusters are anchored to amino acids along a group of proteins that collectively dangle on the matrix side of the inner mitochondrial membrane (see figure 1).
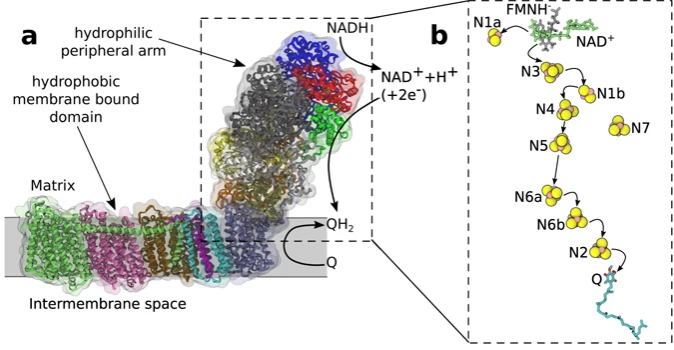
Figure 1: Respiratory Complex I
Image credit: Scientific Reports2
Iron-Sulfur Clusters Add Complexity
These clusters are named based on how many iron and sulfur atoms are present. For example, a FeS cluster called [3Fe-4S] has three iron atoms and four sulfur atoms as part of the core (see figure 2). These clusters may be connected to sulfur-containing amino acids like methionine and cysteine, but those sulfur atoms are not counted as part of the cluster. The clusters are classified as mixed-valence systems, meaning that some of the iron atoms may have a +2 charge and others have a +3 charge. The sulfur atoms have a -2 charge. These differing features lead to many different combinations for the Fe-S clusters, each with varying overall charges and reduction potentials.
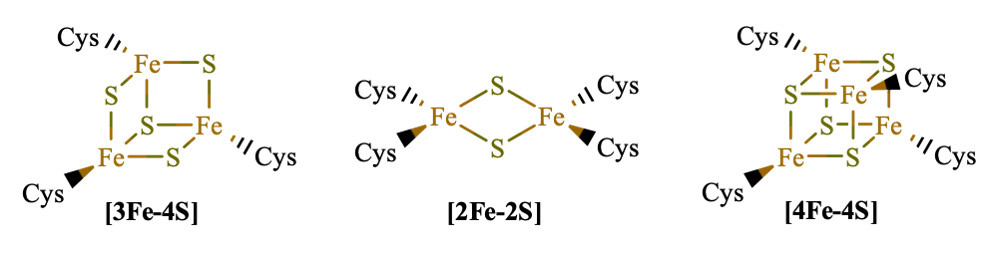
Figure 2: Examples of Iron-Sulfur Clusters
Image credit: Marvin, Chemaxon3
The way the cluster is connected to the larger protein adds variation and fine-tuning. This design feature is critical. If there were only a single metal atom present rather than a cluster of multiple atoms, the reduction potential would not vary at all and electron transport would reach an energetically unfavorable end.
The iron-sulfur clusters are present in three of the five respiratory complexes in the ETC. Respiratory complex I has eight different Fe-S clusters that facilitate multiple electron transfers; however, the total change in potential from start to finish is very small, around 0.4 V.4 For each of these short electron passes, the change in potential must be energetically favorable, from a cluster with a lower electron affinity to one with a higher electron affinity. Further design can be seen in the very small amounts of energy required by the proteins to shift bond distances and angles to accommodate the changes in oxidation state. If this reorganizational energy were too great, the electron transfer would be unfavorable and not occur. Imagine a 300-pound lineman having to contort his upper body 180 degrees to catch the ball.
A byproduct of each of these individual transfers is the release of a very small amount of energy that complexes can use to accomplish the other job of pumping protons across the membrane. This proton gradient is what’s used to help manufacture ATP, the cell’s energy currency. The design of multiple, coupled redox reactions, each releasing a small amount of energy, is much more efficient than having only one reaction that releases a lot of energy; most of which would be lost as heat.5 The overall thermodynamic efficiency of the process is thought to be around 70%. This is not too bad considering that an automobile engine is less than 30% efficient.6
Trick Play or Elegant Design?
The movement of electrons down the ETC seems just as improbable as the game-winning play down the field. Electrons have to move in the right direction (energetically), and the transfers need to be across a short distance, usually around 15 Å or less.7 If the distance is much more than this, the kinetics, or rate of reaction really slows down. Finally, “angry defenders,” that is, reactive species in the cell need to be kept away by the protein matrix to keep the electrons from getting absorbed by the wrong reactant.
The more articles one reads from the scientific literature regarding the ETC, the more one encounters terms like “design,” “wired-together,” “improbable,” and “evolution-perfected,” to describe how truly exceptional and surprising these processes are. Cellular respiration appears to be a very well-designed play. While many scientists try to avoid invoking the designs and actions of a Creator in their descriptions, the harder they try, the more visible the hand of God becomes.
Endnotes
- R. A. Marcus, “On the Theory of Oxidation-Reduction Reactions Involving Electron Transfer,” Journal of Chemical Physics 24, no. 5(May 1956): 966–978, doi.org/10.1063/1.1742723.
- Daniel R. Martin and Dmitry V. Matyushov, “Electron-Transfer Chain in Respiratory Complex I,” Scientific Reports 7 (July 14, 2017): 5495, doi.org/10.1038/s41598-017-05779-y.
- Marvin was used for drawing chemical structures: Marvin 22.14, Chemaxon (chemaxon.com).
- Marina Verkhovskaya and Dmitry A. Bloch, “Energy-Converting Respiratory Complex I: On the Way to the Molecular Mechanism of the Proton Pump,” International Journal of Biochemistry & Cell Biology 45, no. 2 (February 2013): 491–511, doi.org/10.1016/j.biocel.2012.08.024.
- Fazale Rana, “Molecular Logic of the Electron Transport Chain Supports Creation,” The Cell’s Design (blog), Reasons to Believe, February 27, 2019.
- Donald Voet and Judith G. Voet, Biochemistry, 4th ed. (Hoboken, NJ: Wiley, 2011), 829.
- Martin and Matyushov, “Electron-Transfer Chain in Respiratory Complex I.”





